

Стафилококки - обычные обитатели микробиоты носа и кожи, но при этом они легко проникают в соседнюю ротовую полость и дыхательные пути.
В результате их часто выделяют из полости рта и околоротовой области. Изоляты, выделенные из слизистой оболочки полости рта, включают штаммы Staphylococcus aureus с множественной лекарственной резистентностью и кожных комменсалов, таких как Staphylococcus epidermidis и Staphylococcus hominis. Присутствие на этих слизистых оболочках S. aureus и S. epidermidis, связанное с перекрестными инфекциями, делает периоральные области частыми источниками носительства и инфицирования стафилококками.
Отсутствие у этих бактерий механизмов (например, пили и жгутиков) для активной подвижности препятствует рассеиванию путем плавания, а также поверхностного движения или роения. Тем не менее, возможно их прикрепление к жгутиковым подвижным бактериям, таким как Pseudomonas aeruginosa, и совместное рассеивание. Лабораторные исследования показывают, что некоторые штаммы S. aureus и S. epidermidis также способны рассеиваться на высокогидратированных поверхностях. Однако мало что известно о биологическом значении такого поведения и о том, позволяют ли активные клеточные процессы контролировать дисперсию.
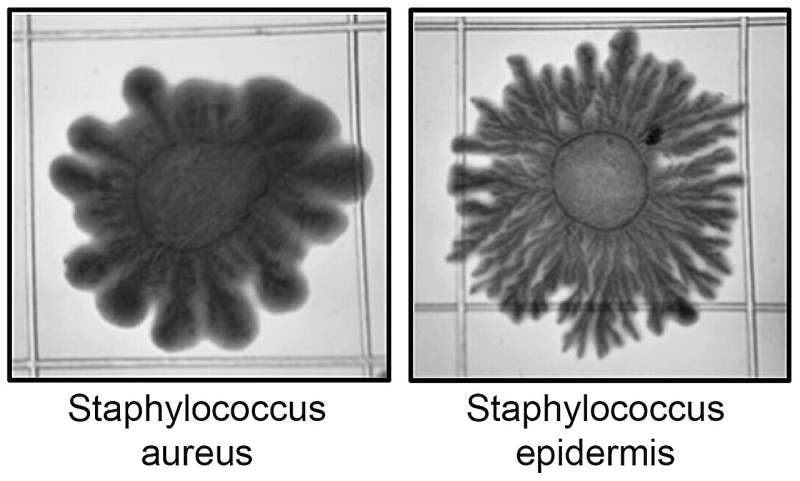
Некоторые штаммы S. aureus могут пассивно распространять свои колонии на мягких агаровых поверхностях при условии, что среда достаточно гидратирована. Например, высокогидратированная агаровая среда [0,24 % (масс./об.) агара, время высыхания 20 мин] способствовала пассивному распространению многих штаммов S. aureus со скоростью до 100 мкм/мин после точечной инокуляции при высокой плотности клеток. Фенотип распространения (с разветвленными ветвями или дендритами) и скорость разрастания колоний были специфичны для каждого штамма и зависели от синтеза и химической модификации тейхоевых кислот клеточной поверхности стафилококков.
Небольшое снижение гидратации среды, например, умеренное увеличение времени высушивания в течение 20 мин, отменяло распространение колоний, что соответствует пассивному способу рассеивания, опосредованному переносом воды. Аналогичные фенотипы колоний были также отмечены на агаровых средах при более высокой концентрации агара [0,34% (масс./об.)], но более коротком (10 мин) времени высушивания. В этих условиях некоторые штаммы S. aureus рассеивались от края колонии в виде бесформенных шлейфов («комет»), которые со временем разрастались в дендритоподобные ветви.
Эти данные подтвердили предположение о том, что вода в среде уменьшает поверхностное трение и способствует распространению клеток из растущих колоний наружу. Также были выдвинуты предположения об активных формах рассеивания, особенно при высокой плотности клеток, которая запускает секрецию поверхностно-активных и смазывающих фенолрастворимых модулинов (PSMs).
Продукция PSM у S. aureus регулируется двухкомпонентной системой кворум-сенсинга Agr (гистидин-киназа AgrC и регулятор ответа AgrA). Фосфорилирование регулятора ответа AgrA при высокой плотности клеток активирует синтез и секрецию PSM, включая два пептида (PSMα3 и PSMγ) с сурфактантной активностью, необходимой для смазывания клеток, движущихся по полутвердой поверхности. Близкий родственник S. aureus, S. epidermidis, имеет отдельную систему agr для регуляции кворум-сенсинга PSMs и также может распространяться по влажной поверхности с помощью механизма, называемого «дартинг», хотя и с гораздо меньшей скоростью (6 мкм/мин), чем S. aureus.
В то время как для распространения колоний S. aureus требуется смазка водой в среде, чтобы уменьшить силы трения между клетками и поверхностью, распространение S. epidermidis происходит в результате «выброса клеток из капсулированного агрегата». Агрегаты S. aureus также могут отделяться и "выкатываться" из микроколоний. Во всех случаях для рассеивания стафилококков на влажных поверхностях требовалась высокая плотность клеток. Кожный комменсал S. hominis также кодирует системы кворум-сенсинга, но не имеет большинства генов для синтеза PSM. Например, лабораторный штамм AH5009 продуцирует только детектируемые уровни PSMβ1 и делает это при определенных лабораторных условиях.
Корреляция между гидратацией среды, секрецией смазывающих PSM и перемещением стафилококков на полутвердых поверхностях агара побудила нас изучить роль, которую могут играть поверхностно-активные, гидратирующие муцины в рассеивании стафилококков по слизистым, особенно обитающих в околоротовой области. Слизистые оболочки человека обычно покрыты слоем слизи, в 104-106 раз более вязкой, чем вода. Контролируемая секреция различных муциновых гликопротеинов нижележащим эпителием регулирует гидратацию, смазку и вязкоупругие свойства слизистого слоя на разных участках тела.
Высокогидратирующие, гелеобразующие муцины, такие как MUC5AC и MUC5B, особенно многочисленны в околоротовой полости и прилегающем слое слизи дыхательных путей, что делает эти слизистые подходящей средой для поверхностного рассеивания потенциально патогенных околоротовых бактерий в нижние дыхательные пути. Действительно, жгутиковые бактерии, такие как P. aeruginosa, плавают в гидратированной среде агара для определения подвижности (Motility Agar) [0,3% (масс./об.)], но не могут перемещаться по его поверхности (роение), пока концентрация агара не увеличится до 0,5% (масс./об.).
Однако жгутиковые клетки используют присутствие всего 0,05 % (масс./об.) гликопротеинов муцина на агаре для перемещения («серфинга») по поверхности агара. Подвижность серфинга у P. aeruginosa регулируется зондированием кворума, но у других серфингирующих видов она не зависит от плотности клеток. Эти различия в контроле плотности клеток согласуются с конвергентным адаптивным ответом для поверхностного рассеивания, облегчаемого муциновой смазкой.
Чтобы проверить аналогичное влияние муцинов на рассеивание периоральных стафилококков, мы изучили фенотипы рассеивания стафилококков, ранее выделенных нами из периоральной слизистой здоровых молодых людей, и сравнили их поведение с тем, которое демонстрируют близкородственные бактерии лабораторных штаммов. Мы провели анализ агаровых чашек для выявления пассивного (распространение колонии наружу) и активного (ветвление или образование дендритов) режимов рассеивания, индуцированных гидратирующими муцинами, обладающими поверхностно-активными свойствами.
Результаты этого исследования подчеркивают стимулирующее влияние смазывающих муцинов на распространение колоний S. aureus и S. epidermidis даже при низкой плотности клеток. Кроме того, наши результаты подчеркивают ключевую роль системы agr-кворум-сенсинга в модуляции секреции PSM и ускорении дендритной экспансии при высокой плотности клеток. Это, наряду с широко распространенной способностью стафилококков использовать муцин в качестве источника углерода и энергии, дает этим видам стафилококков конкурентное преимущество при распространении по слизистой оболочке в периоральных областях и колонизации дыхательных путей человека, где они могут вызывать перекрестные инфекции.
Полученные результаты свидетельствуют о взаимодополняющей роли смазывающих муцинов и стафилококковых PSM в активном рассеивании потенциально патогенных штаммов из периоральной в респираторную слизистую, где в изобилии присутствуют гелеобразующие, гидратирующие муцины. Они также подчеркивают влияние межвидовых взаимодействий на совместное рассеивание S. aureus с другими периоральными бактериями, что повышает риск развития полимикробных инфекций и тяжесть клинических исходов.









