


Прокариотические одноклеточные организмы, предки современных бактерий и архей, самая древняя форма жизни на нашей планете, впервые появившаяся примерно 3,5 миллиарда лет назад.
Первые эукариотические клетки появились примерно 1-1,5 миллиарда лет спустя. Однако с тех пор эукариотические клетки диверсифицировались во множество сложных многоклеточных организмов, которые мы видим вокруг себя каждый день - грибы, растения и животные, в то время как прокариоты оставались исключительно одноклеточными. За всю историю существования жизни на Земле многоклеточные морфологии развивались более 50 раз, причем как в про-, так и в эукариотических линиях. Действительно, некоторые прокариоты достигли версий многоклеточности - например, некоторые фотосинтезирующие цианобактерии разделены на 2 типа клеток, где один тип клеток осуществляет фотосинтез, а другой фиксирует азот. А миксобактерии формируют многоклеточные тела как часть своего сложного жизненного цикла.
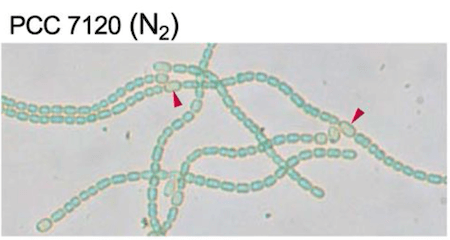
Многоклеточная цианобактерия Anabaena демонстрирует разделение труда на фотосинтезирующие клетки (зеленые) и азотфиксирующие гетероцисты (наконечники стрелок).
Источник: Velázquez-Suárez C., et al./mSphere, 2020
Однако, несмотря на то, что некоторые прокариоты, ведя многоклеточный образ жизни, приблизились к сложной многоклеточности, ни одна прокариотическая линия не достигла сложной многоклеточности так, как это сделали эукариоты - несколько видов клеток работают вместе и организуются в более крупный организм. Действительно, сложная многоклеточность развивалась всего 5 раз, и всегда в пределах эукариот.
Это наблюдение лежит в основе фундаментального вопроса эволюционной биологии: хотя прокариотические клетки имеют сложные клеточные структуры и поведение и у них было достаточно времени для развития сложной многоклеточности, они так и не сделали этого - почему?
Одна из теорий, почему только у эукариот развилась сложная многоклеточность, заключается в том, что только у эукариот развились свойства, необходимые для сложной многоклеточности. Эукариотические клетки отличаются от прокариотических по многим параметрам - они крупнее, имеют большее количество различных органелл и упакованное ядро. Разнообразие форм и функций эукариотических клеток также поражает воображение по сравнению с прокариотами. К ним относятся растительные клетки с жесткими стенками, вытянутые, шиповидные нейроны и реснитчатые клетки, выстилающие стенки нашего кишечника.
Для создания разнообразия форм и функций, наблюдаемого в эукариотических клетках, требуются особые механизмы - в частности, сложные средства генетической регуляции. В процессе развития клеткам должны быть отведены определенные роли. У эукариот ДНК в клетках плотно обернута вокруг белков, называемых гистонами, которые, в свою очередь, упакованы вместе в структуру, известную как хроматин. Считается, что именно такие механизмы регуляции дали эукариотам преимущество при эволюции в сложные многоклеточные организмы.
Модификации гистоновых белков регулируют экспрессию генов во время развития у всех животных, включая самых древних - губок. В 2017 году исследователи из Квинслендского университета в Австралии показали, что один из видов губок обладает чрезвычайно древним регуляторным механизмом, который встречается и у других животных, эволюционировавших гораздо позже. Губка Amphimedon queenslandica морфологически очень проста. Тем не менее, исследователи обнаружили регуляторные системы, которые важны для развития других, более сложных с морфологической точки зрения животных. Одна из них называется Polycomb Repressive Complex 2 (PRC2) - набор белков, которые могут изменять гистоны, чтобы заставить "замолчать" участки хроматина. PRC могут помочь клеткам достичь определенной формы и придерживаться ее, развиваясь в один и тот же вид клеток на протяжении всех циклов деления, что могло способствовать эволюции сложных многоклеточных организмов.
Расширяя свой геном вы расширяете свою морфологию
Однако клеточные свойства - это еще не все. Новое исследование показало, что эволюция сложных многоклеточных организмов могла быть обусловлена невидимой, но мощной силой - генетическим дрейфом. Каждая популяция состоит из особей одного вида, но с вариантами определенных генов. Частота встречаемости различных вариантов генов может меняться под действием отбора, а также в результате случайного стечения обстоятельств. Генетический дрейф включает эти случайные изменения, влияние которых особенно сильно в небольших популяциях с меньшим эффективным размером популяции - числом особей, которые производят следующее поколение.
По мере того как отдельные клетки разделяются на группы клеток в процессе эволюции сложных многоклеточных организмов, размер популяции уменьшается, усиливая эффект генетического дрейфа. Важно отметить, что про- и эукариоты по-разному реагируют на дрейф. Геномы прокариот часто сокращаются перед дрейфом, отчасти из-за встроенной предрасположенности к удалению генов, в то время как у эукариот все происходит наоборот. У эукариот дрейф часто приводит к расширению генома, когда в геном добавляются целые участки ДНК. Прокариоты также могут накапливать гены в результате дрейфа, но обычно это псевдогены - участки ДНК, которые выглядят как гены, но на самом деле не кодируют белок.
Расширение генома может быть сильным фактором эволюции, поскольку оно расширяет генетическую песочницу, в которой можно поиграть в эволюционные новшества. Исследователи использовали в качестве примера цианобактерии - группу бактерий, у некоторых представителей которой простая многоклеточность развилась благодаря набору ключевых адаптаций. Они обнаружили сильную положительную корреляцию между количеством многоклеточных адаптаций и размером генома у почти 200 видов цианобактерий, что говорит о том, что большие геномы важны для многоклеточности у цианобактерий.
Помимо сильной положительной корреляции между многоклеточными адаптациями и размером генома, исследователи также обнаружили, что у многоклеточных цианобактерий более чем в два раза больше псевдогенов по сравнению с одноклеточными видами. Это свидетельствует о том, что их геномы, вероятно, подвергаются эрозии, вызванной дрейфом, в результате многоклеточной адаптации, что также ожидаемо для прокариотических организмов, таких как цианобактерии.
Хотя было предложено множество гипотез, до сих пор не совсем ясно, почему только у эукариот развилась сложная многоклеточность. Кроме того, неизвестно, является ли эрозия генома в результате дрейфа непреодолимым препятствием для эволюции многоклеточности у прокариот. В целом, мы все еще далеки от фундаментального понимания того, как и почему сложная многоклеточность развилась у эукариот, и какие особенности были наиболее важны для основных морфологических переходов. Однако эти вопросы остаются интересными для эволюционной биологии, поскольку ответы на них могут дать нам возможность заглянуть в события, предшествовавшие нашему собственному возникновению сотни миллионов лет назад.









